
| The occurrence and causes of abrupt transitions, thresholds, or regime shifts between ecosystem states are of great concern and the likelihood of such transitions is increasing for many ecological systems. General understanding of abrupt transitions has been advanced by theory, but hindered by the lack of a common, accessible, and data-driven approach to characterizing them. We apply such an approach to 30–60 years of data on environmental drivers, biological responses, and associated evidence from pelagic ocean, coastal benthic, polar marine, and semi-arid grassland ecosystems. Our analyses revealed one case in which the response (krill abundance) linearly tracked abrupt changes in the driver (Pacific Decadal Oscillation), but abrupt transitions detected in the three other cases (sea cucumber abundance, penguin abundance, and black grama grass production) exhibited benthic relationships with drivers (wave intensity, sea-ice duration, and amounts of monsoonal rainfall, respectively) through a variety of response mechanisms. The use of a common approach across these case studies illustrates that: the utility of leading indicators is often limited and can depend on the abruptness of a transition relative to the lifespan of responsive organisms and observation intervals; information on spatiotemporal context is useful for comparing transitions; and ancillary information from associated experiments and observations aids interpretation of response-driver relationships. The understanding of abrupt transitions offered by this approach provides information that can be used to manage state changes and underscores the utility of long-term observations in multiple sentinel sites across a variety of ecosystems. |
|
|
Introduction Many ecological systems can exist in two or more states that differ in abundance or composition of species, rates of ecological processes, and ecosystem services provided by them (Beisner et al. 2003, Suding et al. 2004). Smooth, gradual transitions between ecosystem states are unremarkable, occurring during succession or as ecosystems track gradually changing environmental conditions. In contrast, abrupt transitions between ecosystem states are typically unexpected and can have wide-ranging, negative impacts. Abrupt transitions happen either when the gradually changing environment passes a critical point or when discrete perturbations cause sudden changes in underlying environmental drivers. Abrupt and irreversible transitions are forecast to increase as climatic changes and depletion of natural resources both accelerate (Millennium Ecosystem Assessment 2005, Fagre et al. 2009). Such forecasting, however, is difficult because there are many different causes of state changes (Hastings and Wysham 2010) and because existing approaches demand far more data than are normally available (Carpenter et al. 2011). Managing state changes is as difficult as forecasting them. When environmental changes are not severe, or when organisms with short lifespans and generation times rapidly track environmental drivers, some state changes can be reversed in relatively short periods of time (≤50 years) if drivers are returned to pre-change conditions or perturbations are eliminated (Jones and Schmitz 2009). In other cases, environmental change can result in state changes that persist long after environmental drivers have returned to earlier conditions. The persistence of these socalled ‘‘ecological thresholds’’, ‘‘regime shifts’’, ‘‘phase shifts’’, or ‘‘catastrophes’’ (Hughes 1994, Scheffer et al. 2001, Groffman et al. 2006) is caused by time-lags in the responses of biological systems to environmental change (hysteresis), differences in the relationships between state variables and environmental drivers before and after the state change, or the appearance of novel feedbacks among state variables and drivers that reinforce the new state (Scheffer et al. 2001, Lindig-Cisneros et al. 2003, Briske et al. 2006, Suding and Hobbs 2009). The development of management strategies to mitigate abrupt transitions requires strong linkages among theory, data, and case studies, but there is little guidance available for using historical or ongoing studies to detect or respond to abrupt transitions. There is confusion and disagreement about what changes constitute transitions (Rudnick and Davis 2003, Schroder et al. 2005) and a limited understanding of ecological mechanisms causing them (Brown and Archer 1999, Collie |
et al. 2004). Empiricists disagree about how to best gather and interpret relevant data (Petraitis and Latham 1999, Bertness et al. 2002, Schroder et al. 2005), while theoreticians develop leading indicators of abrupt transitions that demand large amounts of data (Carpenter and Brock 2006, Biggs et al. 2009, Contamin and Ellison 2009). There is little clarity regarding the use of existing data and the design of future studies to detect and mitigate undesired state changes (Bestelmeyer 2006, Groffman et al. 2006). A common, systematic approach to analyzing state changes could allow ecologists to marshal a large body of useful data and detailed knowledge to help society better understand and, ultimately, manage abrupt transitions. Here, we illustrate a general, data-based, and mechanism-centered analysis of abrupt transitions using four datasets from the US Long-Term Ecological Research (LTER) program on pelagic ocean, coastal benthic, polar marine, and semi-arid terrestrial ecosystems. These LTER data include some of the longest time-series available for both causal environmental drivers and biological response variables, and interpretations of associations between the drivers and the response variables are enhanced by experimental and mechanistic studies conducted at the same sites. We first lay out a synthetic framework for describing abrupt transitions and state changes that can be used to compare and contrast among case studies. We then propose a standard analytical approach that provides strong tests for detecting abrupt transitions between states. This approach revealed unexpected results for the pelagic ocean system for which a ‘‘regime shift’’ had been described previously, provided stronger evidence for hypothesized state changes in the coastal benthic ecosystem, and yielded new evidence for state changes in the polar marine and semi-arid terrestrial ecosystems. Our analyses illustrate how to identify and interpret causes of abrupt transitions, and also illustrate limitations common to many datasets used to study abrupt transitions and state changes. We conclude with recommendations for improving ongoing and nascent long-term research programs aimed at detecting and forecasting state changes. A common framework for describing state change |
|
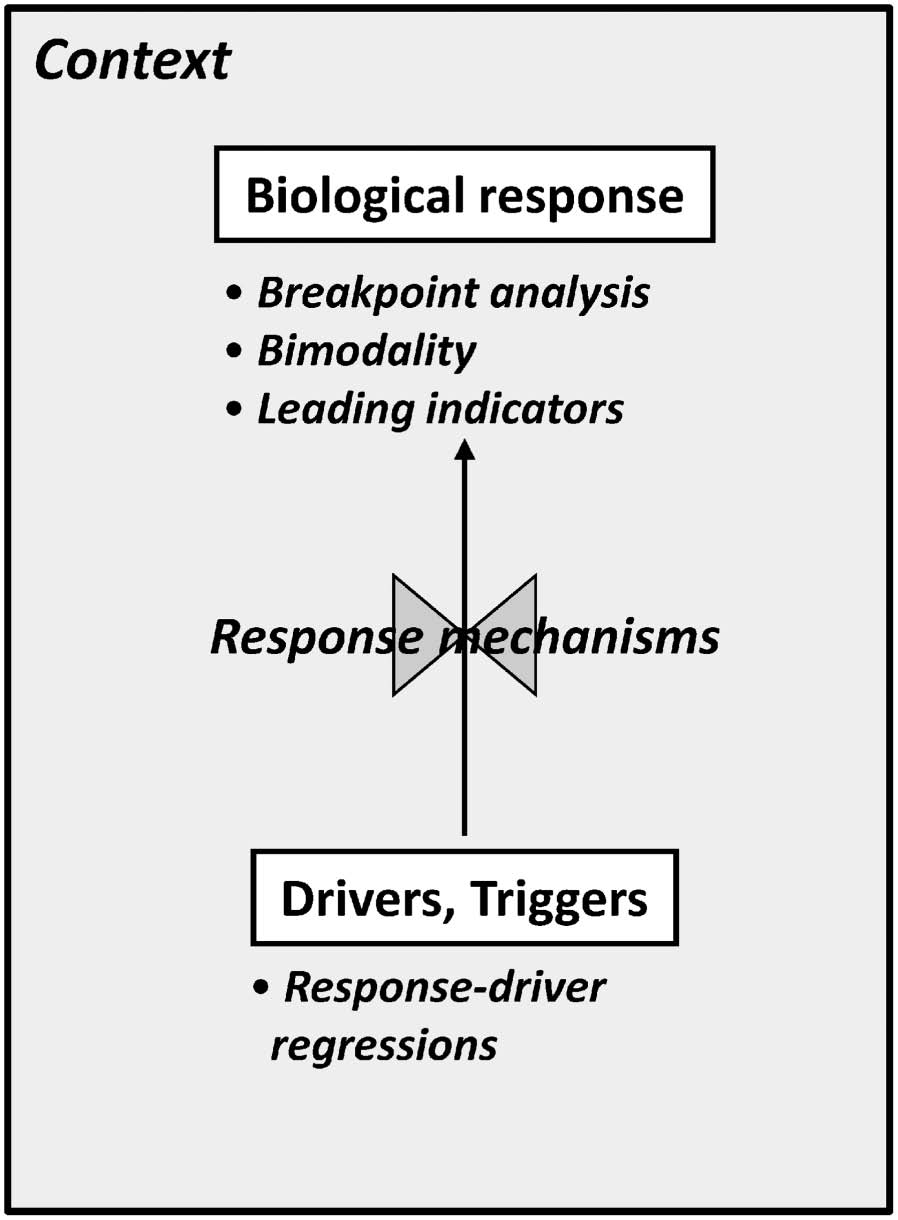
Fig. 1. A conceptual model of the relationships between the elements of abrupt transitions and analytical approaches used to investigate them.
|
|
|
State changes in biological responses are caused directly or indirectly by changes in environmental drivers. Drivers are usually abiotic and include changes in climate (e.g., temperature, precipitation), or land-use (e.g., resource extraction, nutrient input rates). Environmental drivers usually are considered ‘‘slow variables’’ (e.g., Folke et al. 2004, Carpenter and Brock 2006) because they typically change much more slowly than biological response variables (Fig. 2A). The textbook example of a slow environmental driver leading to a state change is long-term phosphorus input leading to an abrupt shift from oligotropic (clear blue) to eutrophic (muddy green) lakes (Carpenter and Brock 2006). Drivers can also change abruptly, however, with dramatic effects. Triggers (a.k.a. pulse disturbances) are either abrupt shifts in drivers or singular events, such as droughts, hurricanes, disease outbreaks, invasive species introductions, or fire, that directly affect biological responses (Suding and Hobbs 2009). State changes often are caused by interactions among multiple drivers and triggers (Nystrom et al. 2000, Breshears et al. 2005). Whereas drivers are typically presented as time series concurrent with biological responses (Fig. 2A), triggers are discrete events in time or relatively short, discrete sections of a time series (e.g., an El Niño period, Holmgren et al. 2006). Biological responses (a.k.a. response variables or state variables) are used to recognize alternative states |
(Mantua 2004, Schroder et al. 2005, Andersen et al. 2009). Response variables are especially important because they usually can be measured or monitored easily, and persistent changes in their mean or increases in their variance are used as indicators of state changes. Like environmental drivers, biological response variables typically are represented as time series of the abundance or biomass of individual species or suites of trophically similar species (Daskalov et al. 2007; Fig. 2A). Response mechanisms describe how drivers and triggers interact and affect responses (Fig. 1). Of particular importance are changes in relationships between drivers and responses caused by positive feedbacks between them that amplify changes in both drivers and responses and reinforce alternative states (Rietkerk et al. 2004). Positive feedbacks often involve complex chains of interactions involving biological and physical processes, including Allee effects (Bourbeau-Lemieux et al. 2011), trophic cascades (Carpenter et al. 1999, Carpenter et al. 2011), habitat fragmentation and extinction cascades (Swift and Hannon 2010, He and Hubbell 2011), land surface-climate feedbacks (Foley et al. 2003, Cook et al. 2009), or spreading desertification (Peters et al. 2004). Data on response mechanisms are derived most frequently from manipulative experiments, natural history observations, and expert knowledge (Choy et al. 2009). |
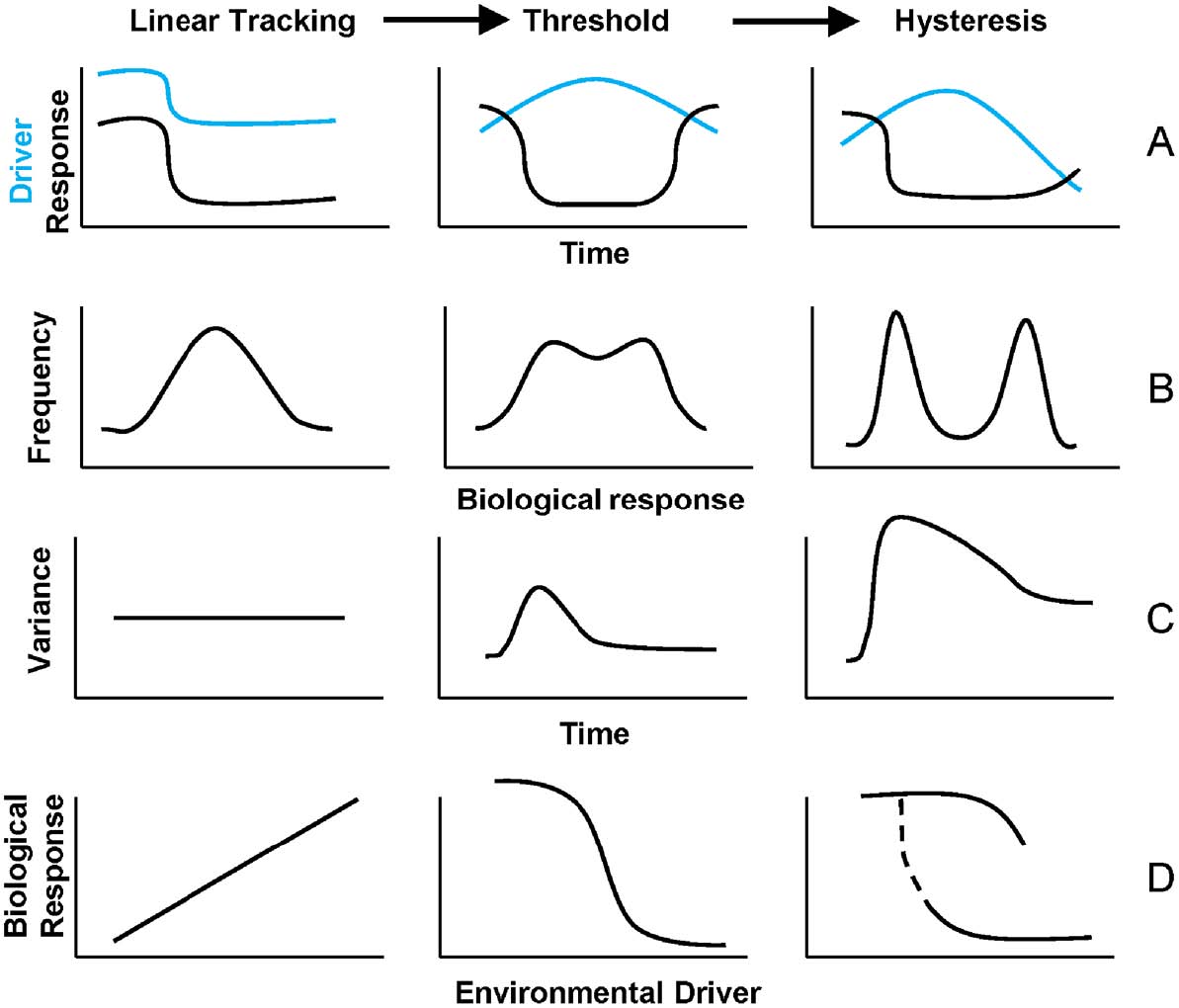 |
Fig. 2. Three classes of driver-response relationships and analytical indicators of transitions and state changes. The top row (A) illustrates time series of driver and response variables in linear tracking, threshold, and hysteretic systems. The second row (B) illustrates that the frequency distribution of the observations should shift from unimodal to bimodal when a threshold or hysteretic change occurs. The third row (C) illustrates how one leading indicator, the variance of the time series, should differ among the three classes of driver-response relationships. As the transition becomes more abrupt and the post-transition state becomes more distinctive from the pre-transition state, the variance should become more peaked at the transition point. The bottom row (D) illustrates changes in the driver-response relationships from linear (in the linear tracking class) to nonlinear (in the threshold class) to hysteretic. |
|
Finally, contextual information documents characteristics of the environmental setting that can influence driver-response relationships and that can vary among case studies. For example, lake morphometry (Genkai-Kato and Carpenter 2005), stream channel geometry (Heffernan et al. 2008), soil texture (Bestelmeyer et al. 2006), and distance to source populations (Hughes et al. 1999) result in spatial variation in biological responses to drivers and triggers. Similarly, the timing of disturbance events with respect to seasonal period can determine their effects on biological responses (Nystrom et al. 2000). Understanding spatiotemporal context can help to reconcile differences among case studies illustrating general types of transitions and state changes (e.g., Petraitis et al. 2009). Contextual information also can help translate scientific analyses into meaningful policy recommendations and management interventions (Carpenter et al. 2011). An approach for identifying abrupt transitions and state changes in ecological systems The distinction between linear tracking and threshold responses is whether: the distribution of the biological response variable is unimodal vs. weakly bimodal (Fig. 2B); the variance in the biological response is constant vs. increasing slightly as the environment changes or a trigger occurs (Fig. 2C); and the relationship between the environmental driver and the biological response is linear vs. nonlinear (Fig. 2D). Following the terminology of preceding authors, both linear tracking and threshold responses can be reversed; as the driver returns to its initial (pre-change) value, environmental conditions and biological response variables often track them with at most short time-lags. Note that the threshold in ‘‘threshold response’’ refers to the nonlinear biological response to a change in driver magnitude, rather than irreversibility. In contrast to threshold responses, hysteretic responses result from persistent environmental changes, changes in feedbacks between drivers and response variables, or long time lags in biological responses to drivers. In such systems, even if the environmental driver returns to earlier values, the biological response may not return to its earlier state, or does so only slowly, at a markedly different magnitude of the driver, or along a different path from the one it took to reach its new state (Fig. 2D). The functional form of the relationship between environmental driver(s) and biological response(s) typically differs before and after a state change. |
Results Of This Research A pelagic ocean ecosystem: The six-decade California Cooperative Oceanic Fisheries Investigations (CalCOFI) record revealed that N. simplex generally was rare when the PDO was in the negative phase (anomalously cool waters in the NE Pacific). Abrupt changes of the PDO from negative to positive were tracked by increases in N. simplex abundance and vice versa (Fig. 3A, B). Strong El Niño (1958–60, 1997–98) and La Niña (1998–99) events had positive and negative influences, respectively, on N. simplex abundance that interacted with changes in the PDO to accentuate abrupt changes or interrupt relationships with PDO (Appendix). Although the warm phase between 1977–1998 was a period of consistently high abundance of N. simplex relative to the time period before and after, our data indicate that contrary to previous work (e.g., deYoung et al. 2008) this should not be considered a different ecological ‘‘regime’’ and hysteresis is not indicated. The frequency distribution of abundances were unimodal and, most definitively, the relationship between N. simplex abundance (response variable) and the PDO (driver) varied linearly with the variations in the PDO and was identical in both the warm and the cool phases of the PDO. Thus, the California Current System illustrates a case of linear tracking (Hsieh and Ohman 2006), without discrete, definable (or ‘‘preferred’’) system states. Such linear tracking may be common in shortlived organisms that can quickly and closely track abrupt changes in drivers. |
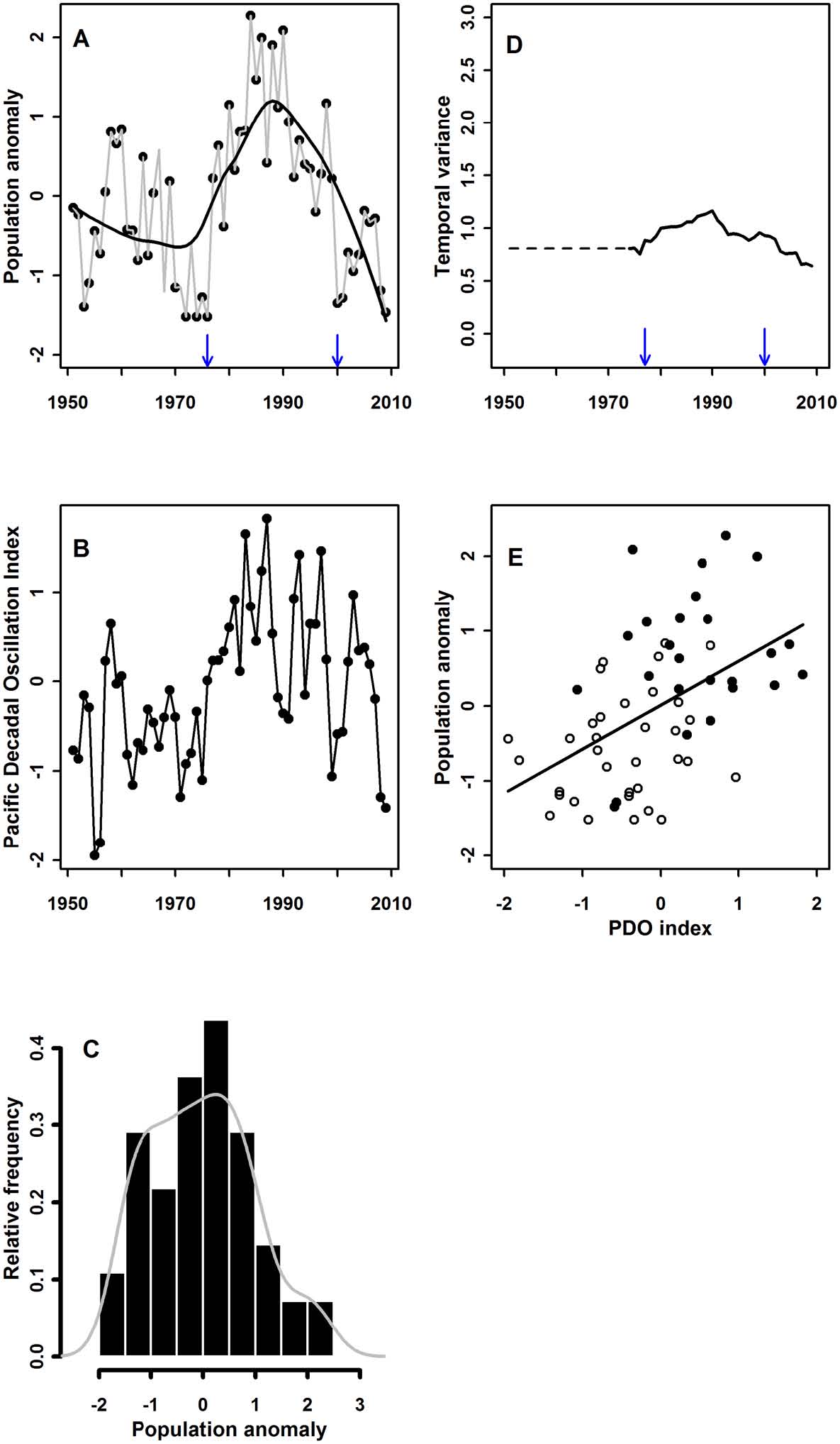 |
Fig. 3. Results of analyses for the California Current System. In the time series of the biological response (A), the observed data are shown as points scaled in standard deviation units, the time series (which includemodeled values) as grey lines connecting the points, and the locally weighted regression (loess) illustrating the temporal pattern as a solid black curve. Breakpoints identified using CUSUM, RSS, and BIC are identified with arrows on the x-axis. The potential alternate state begins the year after the breakpoint. In the time series of the driver (B), the observed data are shown as points and the time series as black lines connecting the points. There are nomissing values in the time series of drivers in Figs. 3–6. The frequency distributions (C) are equivalently scaled across Figs. 3–6, and all bins are the same width (0.5 SDunits). The probability density function of the observations is overlain on the histogram. Similarly, the time series of variance (D) are all equivalently scaled across Figs. 3–6, and breakpoints again are identified. |
|
A coastal benthic ecosystem: Time-series data from nine sites spread along a 5-km stretch of coastline on the north shore of Santa Cruz Island illustrate the mechanisms of abrupt sea cucumber-to-macroalgae transitions (Fig. 4A; see also Rassweiler et al. 2010). The frequency distribution of annual sea cucumber abundance data revealed |
evidence of bimodality (Fig. 4C). The first transition from macroalgae to sea cucumber dominance occurred in the late 1980s and was associated with a series of years in which there were few high wave events during winter storms (Fig. 4B). High waves dislodge sea cucumbers from algal beds (Rassweiler et al. 2008), but when winter storms are weak, sea cucumbers competitively displace algae by smothering and killing them. Frequent, strong storms returned after 1995, but the relationship between days of high waves (driver) and sea cucumber abundance (response) disappeared and sea cucumbers continued to dominate the system (Fig. 4E). Consumption of algal spores by abundant sea cucumbers allowed this species to persist in the face of increased wave disturbance (Rassweiler et al. 2008). This relationship switched to yet another low cucumber state when predatory sea stars colonized the system in late 2002 (Appendix). Thus, this case conforms to a hysteresis model in which stabilizing feedbacks conferred resilience with respect to the environmental driver. |
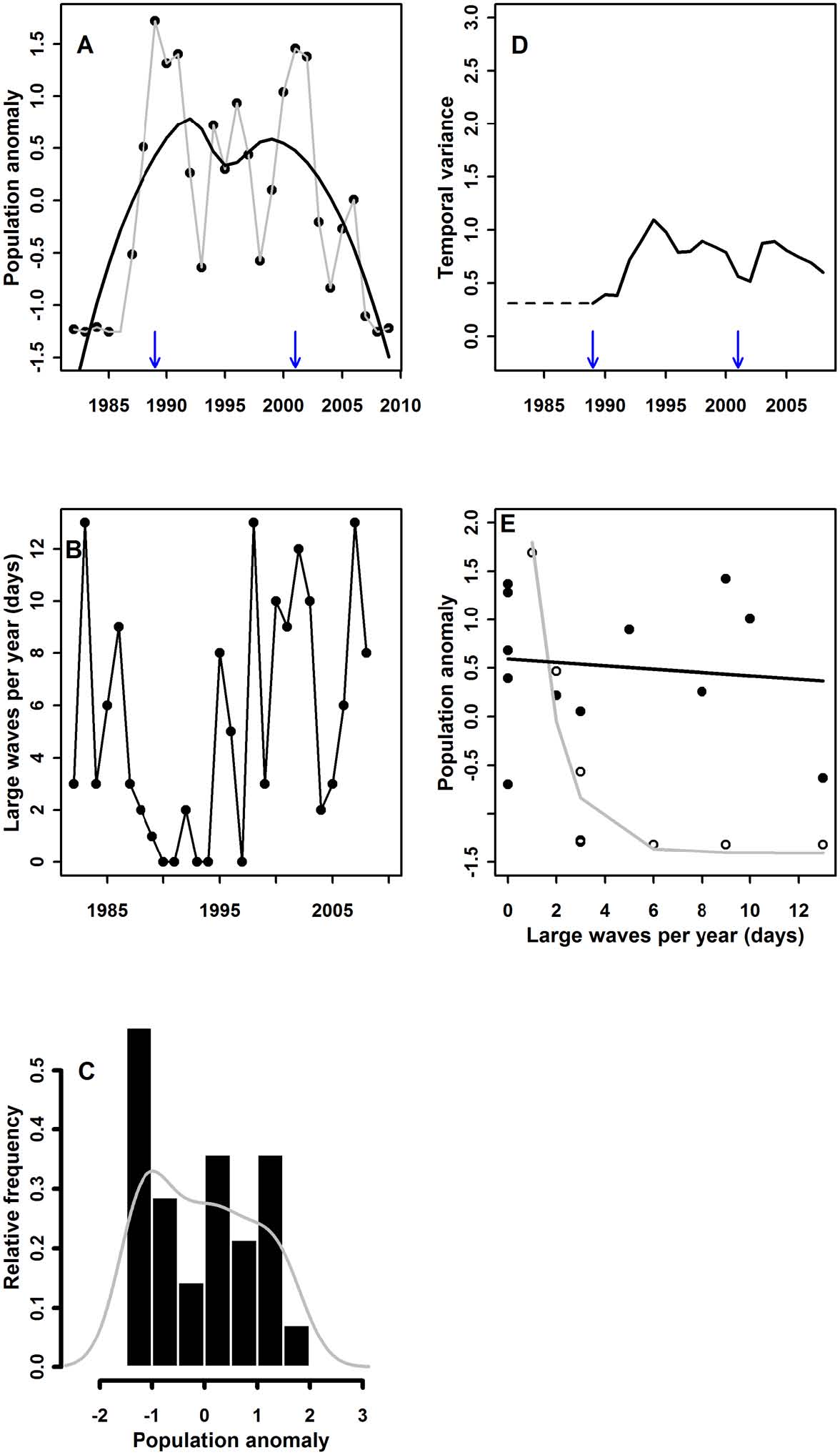 |
Fig. 4. Results of analyses for the Southern California Rocky Reef (A–E), following the same order and rules as for Fig. 3. For the relationship between the driver and response (E), only data from the first and second states are shown. |
|
A polar marine ecosystem: Nearly 40 years of data collected from the Palmer Archipelago near Anvers Island, Antarctica (Appendix: Fig. A2) |
illustrate abrupt declines in the Adélie penguin breeding population beginning in 1993, and abrupt increases in numbers of breeding chinstrap and gentoo penguins beginning in 2004 (Fig. 5A). Bimodality of annual abundance data was not clearly evident (Fig. 5C), nor did temporal variance (Fig. 5D) illustrate dramatic changes before or during the observed population changes. However, analysis of the relationship between the proximate driver (sea-ice duration; Fig. 5B) and Adélie penguin breeding population size revealed that prior to the 1993 breakpoint, the abundance of this species was essentially unresponsive to variation in sea-ice duration, however after 1993 these variables were strongly and positively correlated (Fig. 5E), conforming to the hysteresis model. We did not examine driver-response relationships for chinstrap or gentoo penguins because only five data points on yearly numbers of breeding pairs have been obtained since the 2004 breakpoint. Progressive climate warming resulted in an abrupt transition operating through multiple, cascading ecological drivers and feedbacks, including reduced sea-ice duration, changes in terrestrial snowfall accumulation that affect penguin breeding biology, and feedbacks between Adélie population reductions and predator efficiency (Appendix). |
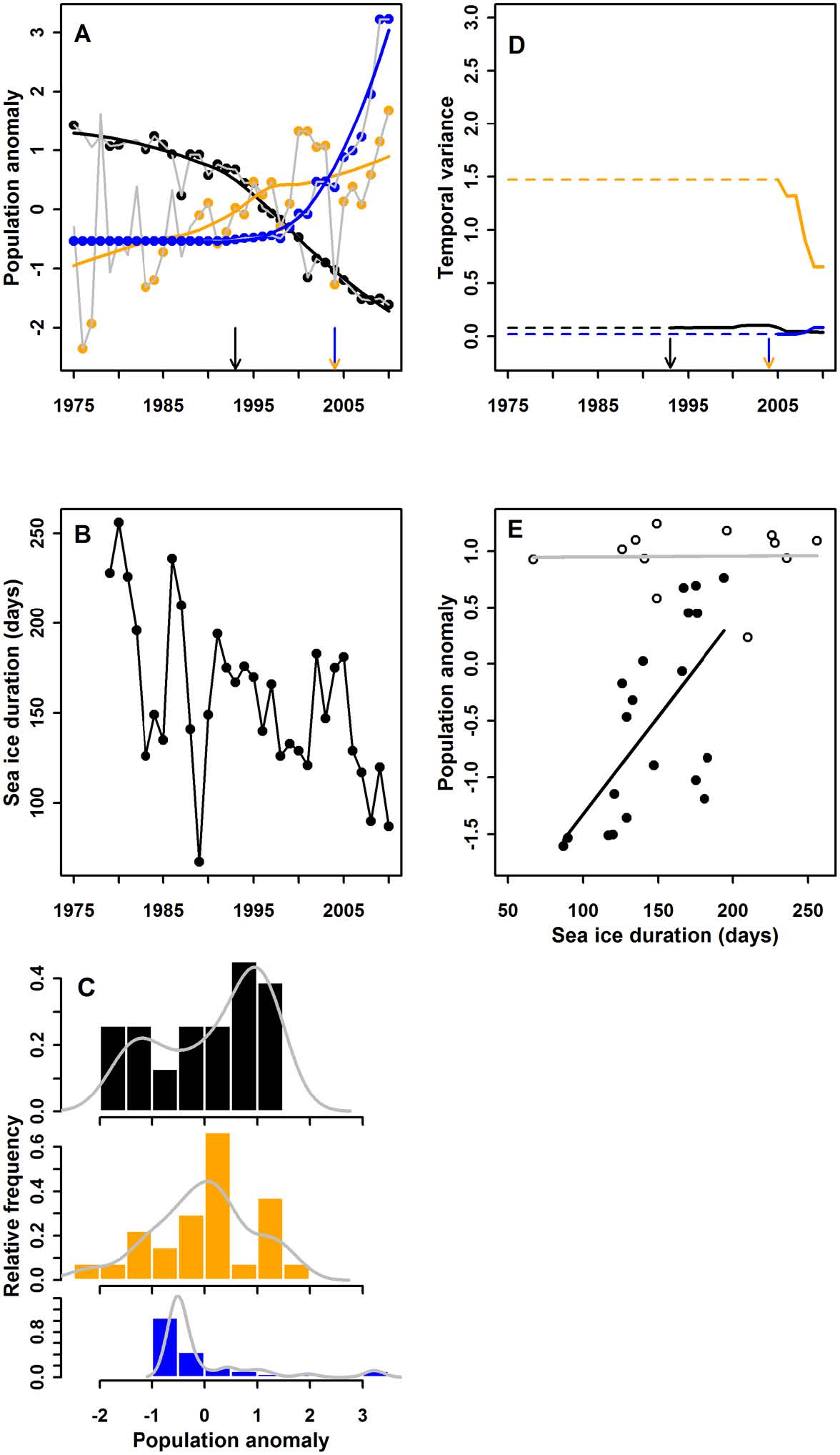 |
Fig. 5. Results of analyses for the Western Antarctic Peninsula (A–E), following the same order and rules as for Fig. 3. The three species of penguins are illustrated in three colors (Adélie penguins in black, chinstrap penguins in orange, gentoo penguins in blue). The relationship between the driver and response is shown only for Adélies because there are too few data for the other species. |
|
A semi-arid grassland ecosystem: Time-series data on annual production of black grama grass collected during the mid-1900s from two pastures in the Jornada Experimental Range near Las Cruces, NM, USA, indicate the start of an abrupt transition in 1948. In that year, there was no black grama production (Fig. 6A), and this lack of production coincided with the onset of a prolonged drought (Fig. 6B). Several lines of evidence suggest that this system conforms to the hysteresis model. First, annual production was bimodal (Fig. 6C), indicating two alternative states. Second, black grama production exhibited an increase in temporal variance during the transition (Fig. 6D) associated with a period of low and variable growing-season (July–September) rainfall |
(Fig. 6B). Third, driver-response regressions show that prior to 1948, black grama production had a positive relationship to growing-season precipitation (Nelson 1934). After 1948, however, this relationship weakened and overall production was low regardless of growing-season rainfall (Fig. 6E). The shift in black grama production was very abrupt, never attaining previous high values after 1950. A positive feedback between soil erosion and low grass cover appears to have precluded grassland recovery after a return to higher levels of precipitation (Appendix). To summarize, we used a common approach to determine if and when abrupt transitions occurred, to evaluate leading indicators that could forecast the transitions, and to match each case to the appropriate class of mechanisms (Figs. 3–6). The timing of abrupt transitions was successfully identified in all four cases. An increase in variance that could serve as a leading indicator was observed only in the Chihuahuan Desert case due to the extreme interannual fluctuation preceding grassland collapse. The linear tracking model was indicated for the California Current case due to unimodality in the distribution of biological response values and linearity in the driver-response relationship. The hysteresis model was indicated in the other three cases due to varying combinations of evidence, including the strong nonlinearity in the driver–response relationship for the West Antarctic Peninsula, and both bimodality of biological responses and nonlinear driver-response relationships in the Southern California Rocky Reef and Chihuahuan Desert cases. In all four cases, plausible response mechanisms supported the classification of the case to the general mechanism. |
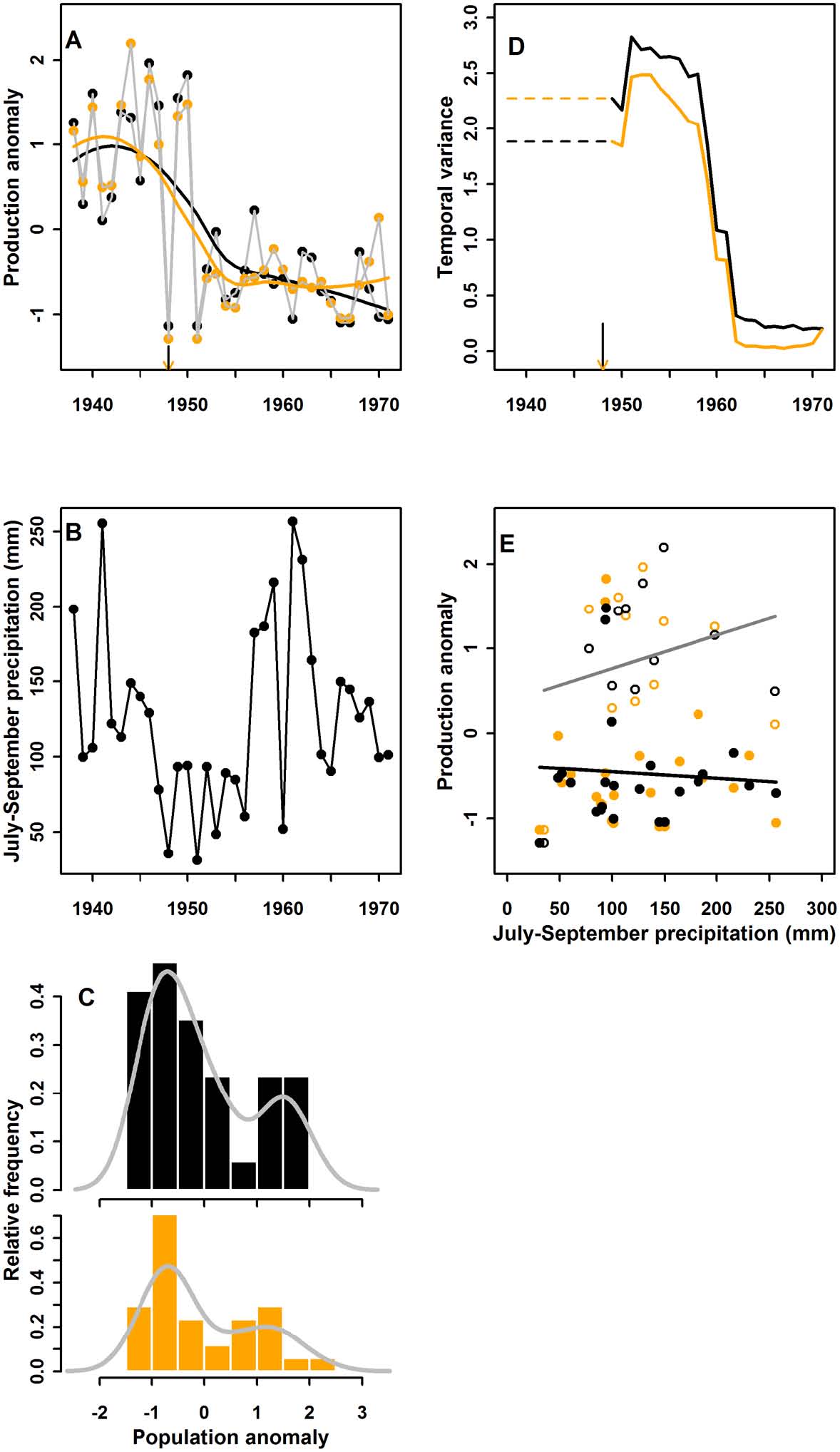 |
Fig. 6. Results of analyses for the northern Chihuahuan Desert (A–E), following the same order and rules as for Fig. 3. The two pastures are illustrated in black (Pasture 2) and orange (Pasture 9). |
| For methods, discussion, and literature cited, see this publication in Ecosphere - Bestelmeyer, B. T., A. M. Ellison,W. R. Fraser, K. B. Gorman, S. J. Holbrook, C. M. Laney, M. D. Ohman, D. P. C. Peters, F. C. Pillsbury, A. Rassweiler, R. J. Schmitt, and S. Sharma. 2011. Analysis of abrupt transitions in ecological systems. Ecosphere 2(12):129. doi:10.1890/ES11-00216.1 |

